Nat Rev Immunol:个性化新抗原肿瘤疫苗面临的挑战
2023-10-11 小药说药 小药说药 发表于上海

在开发发现新抗原并引发有效的新抗原特异性抗肿瘤T细胞免疫的技术方面已经取得了相当大的进展,但疫苗要发挥抗癌功效,仍有许多障碍需要克服。
肿瘤细胞中发生的突变可以产生新的自身抗原表位,称为新表位或新抗原。基于新抗原而非传统TAAs的疫苗有几个优点。首先,新抗原仅由肿瘤细胞表达,因此可以引发真正的肿瘤特异性T细胞反应,从而防止对非肿瘤组织的“脱靶”损伤。第二,新抗原是源于体细胞突变的新表位,它有可能绕过T细胞对自身表位的中心耐受,从而诱导对肿瘤的免疫反应。此外,这些疫苗增强的新抗原特异性T细胞反应持续存在并提供治疗后免疫记忆的潜力,这为长期预防疾病复发提供了可能性。
到目前为止,已有100多项新抗原癌症疫苗的临床试验正在进行或已完成。然而,这些治疗性疫苗试验还没有显示出明显的临床益处,直到最近的两项试验——黑色素瘤的NCT03897881和胰腺癌的NCT04161755——报告了延长的无复发生存期(RFS)。尽管有这些临床疗效的初步证据,但在新抗原癌症疫苗能够充分发挥其临床潜力之前,仍必须解决一些关键挑战。
新抗原疫苗的免疫原性
迄今为止,许多新抗原靶向癌症疫苗已进入临床试验,其中包括基于肽、DNA、RNA、腺病毒和树突状细胞(DC)的疫苗,其中50%以上的试验使用合成长肽(SLP)加佐剂组合。尽管这些疫苗引发了针对癌症相关靶点的T细胞免疫,但仍然存在的一个主要问题是它们的总体免疫原性较低。
通常,在这些试验中引发的新抗原特异性T细胞反应,特别是细胞毒性CD8+T细胞(CTL)反应,在直接活体检查患者外周血时是检测不到的或非常小。在这类研究中,一种常见的做法是报告体外扩增后测量的新抗原特异性T细胞反应。而一般的病原体特异性疫苗,如黄热病减毒疫苗(YFV-17D疫苗)和牛痘病毒活疫苗(Dryvax天花疫苗)都能引发强大的终身免疫,针对单个表位的病毒特异性效应CTL反应约占血液中总CTL的10-20%。所以毫不奇怪,这些新抗原癌症疫苗没能提供足够的临床益处。
目前新抗原疫苗的免疫原性较低,很可能是由于新抗原的内在特征、它们提供的低抗原载量及其次优的免疫刺激能力。此外,在新抗原疫苗临床研究中经常被忽视的一个方面是T辅助细胞2(TH2)免疫的诱导,这种反应主要通过中和抗体来防止病原体感染和传播,但不能促进细胞内病原体的清除。然而,在癌症疫苗开发的情况下,最佳抗肿瘤反应通常涉及新抗原特异性TH1细胞和CTL,但不涉及TH2免疫。TH2细胞在肿瘤环境中的保护作用仍然存在争议,因为它们可以支持更多的抗炎和耐受反应,从而促进肿瘤生长。由于胸腺中T细胞对自身抗原的阴性选择,新抗原特异性T细胞受体(TCRs)的亲和力往往较低,这种低TCR亲和力通常倾向于使免疫偏向TH2型而不是TH1型免疫应答。因此,通过用疫苗佐剂的特定组合进行免疫调节以获得最佳抗肿瘤反应可以避免或克服这些缺点。
新抗原的选择瓶颈
任何疫苗的效力都取决于其抗原的质量和免疫原性,新抗原疫苗也不例外。二代测序(NGS)为个体肿瘤和个体患者提供了前所未有的基因信息。这使得能够通过生物信息学分析和MHC-肽结合预测算法来预测潜在的新抗原肽。目前,已经开发了许多生物信息学方法来预测新抗原肽。然而,这些生物信息学方法的效率仍然存在争议。在一项对13项已发表研究的荟萃分析中,使用的生物信息学预测的新肽只有<2.7%被患者来源的T细胞识别。
另一方面,依赖患者已有的抗肿瘤T细胞免疫来验证肽靶点的缺陷是,肿瘤细胞表达和呈递的一些新肽可能无法在患者体内自发引起T细胞免疫。根据这种选择方法,可能会丢弃在接种疫苗后特异性诱导保护性抗肿瘤反应的新肽。这种新肽可能由肿瘤或APC上的MHC分子低表达或低水平呈递,但如果与强效佐剂结合,它们很可能具有免疫原性。事实上,MHC I通过直接递呈途径(在肿瘤细胞中)或交叉递呈途径(在APC中)呈递的效率显示出对抗原性质的不同要求。例如,基于插入或缺失的肿瘤突变可以导致多肽序列的移码翻译,与野生型对应物几乎没有同源性。随之而来的功能和结构稳定性的缺乏使这些新抗原成为有缺陷的核糖体产物(DRiPs),其是快速降解的靶标,有效地提供直接的MHC I类呈递途径。而交叉递呈已被证明主要依赖于稳定的抗原,而不是DRiPs,这可以解释为什么移码新抗原以及其他蛋白质稳定性降低的突变自身抗原不能充分地启动交叉呈递反应。
突变依赖性新抗原的另一个普遍问题是在原发性癌症中以及在转移性肿瘤中发现的肿瘤内异质性和克隆多样性。肿瘤内的基因多样性表明,并非所有新抗原都是疫苗的良好靶点。优选地,癌症中所有肿瘤细胞携带的主干突变将比分支突变更好地作为疫苗靶点。然而,破译最普遍的肿瘤克隆所携带的真正的干-支突变并非易事。肿瘤的免疫编辑对最具免疫原性的肿瘤抗原施加免疫选择压力,从而使其能够逃避T细胞免疫。疫苗靶向的新抗原也可能发生这种情况,如果使用次优的抗原-佐剂组合,肿瘤的遗传异质性和免疫编辑可能会阻碍新抗原疫苗的效果。表位扩散,即T细胞对疫苗中未包含的抗原的免疫范围的扩大,可能提供一种针对这种逃逸克隆的手段。因此,如何在接种新抗原疫苗后最大限度地扩大表位传播是未来的一个重要挑战。
从以上可以清楚地看出,免疫相关新肽的预测仍然是开发个性化癌症疫苗的主要障碍。确定哪些新肽能够真正呈现在肿瘤表面的MHC I类分子上,哪些能够有效地由DC交叉呈现,仍然是一个挑战。
新抗原肿瘤疫苗的设计
两个经常被忽视的重要问题如下:最佳的新肽表位是什么?新抗原靶向疫苗需要包括多少个表位才能发挥效力?虽然高TCR亲和力的高表达肽足以引发肿瘤损伤和炎症,从而导致表位扩散和肿瘤消除;然而,这可能不会发生在TCR亲和力较低的新抗原中,这些新抗原没有足够的免疫原性来启动T细胞免疫。
因此,有效疫苗所需的新肽的数量可能取决于所包含表位的性质。针对重要驱动突变的高TCR亲和力新肽,即使在肿瘤突变负担较低的肿瘤中也能提供适度的临床益处。然而,由于大多数新肽显示出低TCR亲和力,因此可能需要大量这样的肽来触发才能达到高TCR亲和力抗原的炎症水平和表位扩散。
新肽的长度也是疫苗设计中的一个重要考虑因素。使用可通过肽交换直接负载到MHC I类分子上的短肽可导致在非APC(如T细胞)或非免疫细胞(如成纤维细胞或上皮细胞)的表面上呈递,这可能导致耐受性并促进肿瘤生长。使用依赖于交叉呈递的较长肽可能提供免疫原性优势。原则上,长度编码27个氨基酸的SLP、RNA或DNA可以包含所有理论上可能的MHC I和MHC II类肽,用于14位的单个突变。
佐剂的选择
尽管新肽的选择仍然是疫苗开发的一个重要障碍,但另一个重要问题在于疫苗佐剂的效力。新肽疫苗尤其需要佐剂,因为源自肿瘤突变的新肽通常与自身肽相似,因此TCR亲和力较低,而在没有佐剂的情况下,肽通常缺乏足够的免疫原性。
一些疫苗平台,如病毒、质粒DNA和mRNA疫苗,可以作为其自身的佐剂,因为它们可以通过多种模式识别受体(PRRs)刺激先天免疫,如Toll样受体(TLRs)、RIG-I样受体和cGAS–STING通路。其他平台,如基于肽的疫苗,需要外源佐剂来刺激免疫,这对于诱导强大的CD4+和CD8+T细胞反应至关重要。
在过去的100年里,只有六种佐剂被批准用于人类。这些包括铝佐剂、水包油乳液角鲨烯佐剂MF59、AS01(其含有单磷酰脂质A加皂苷组分QS-21)、AS04(其含有铝佐剂加单磷酰脂A)、AS03(其含有角鲨烯加维生素E)和合成寡脱氧核苷酸CpG。这些佐剂主要设计用于诱导抗体介导的保护,可以诱导不同程度的TH1型和TH2型混合免疫反应。
鉴于新抗原癌症疫苗需要增强T细胞反应的佐剂,一些最有效的T细胞免疫刺激分子如PRR激动剂,越来越多的正被测试用于癌症疫苗设计中的佐剂。CpG-ODNs可以通过刺激TLR9来激活APC,特别是,K3型CpG-ODN是一种诱导体液和细胞免疫的强效佐剂。STING激动剂也可以作为有效的佐剂,通过诱导I型IFNs1扩增抗原特异性CD8+T细胞。重要的是,TH2型免疫应答也通过STING激动剂增强,而K3 CpG-ODNs和STING激动剂的组合可以阻断TH2型反应并协同诱导TH1型免疫反应。
因此,在没有TH2细胞诱导的情况下,鉴定增强疫苗免疫刺激能力以促进TH1细胞和CTL免疫的佐剂,同时避免毒性增加,仍然是新抗原疫苗的总体挑战。
接种疫苗后的挑战
开发有效的个性化新抗原特异性T细胞疫苗可能只是产生临床抗肿瘤疗效的第一步。当使用新抗原疫苗后,为了使有效的T细胞诱导疫苗发挥其完全的抗肿瘤作用,仍然需要克服许多障碍。
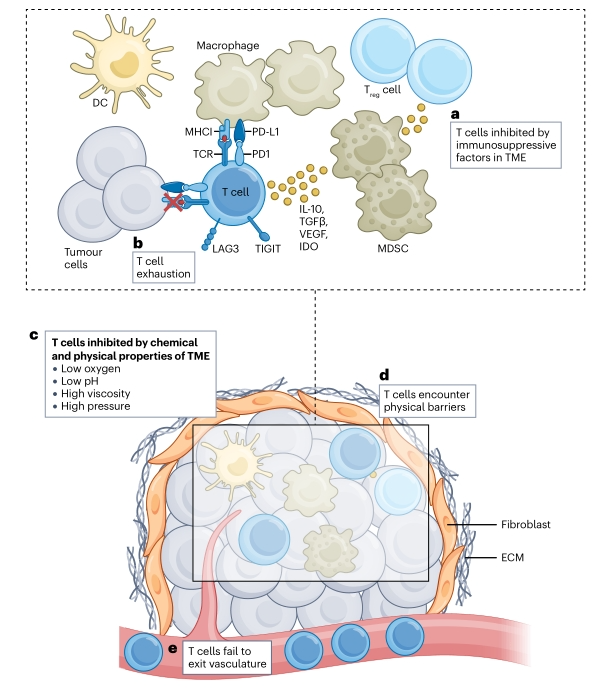
免疫系统的可及性
免疫系统与肿瘤直接相关的两个问题是:T细胞无法识别逃避免疫的肿瘤,以及免疫抑制性TME对T细胞的抑制。
在许多最常见的人类癌症中观察到MHC I类的低表达,这可能使肿瘤对免疫系统具有抵抗力。这种MHC I类下调在分子上是由MHC I级通路上几乎任何地方发生的缺陷引起的。在肿瘤进化水平上,MHC I类下调是T细胞免疫介导的克隆选择的结果,该克隆选择有利于缺乏MHC I型表达的肿瘤克隆逃避T细胞免疫。
上调MHC I的策略可能涉及TLR激动剂、STING激动剂和细胞因子(即TNF、IL-1以及I型和II型干扰素)。化学治疗剂和电离辐射也可以增加肿瘤中MHC I类表面的表达,但这些方式也可以是免疫抑制的,需要良好的平衡。
在TME中,一旦新抗原疫苗诱导的T细胞进入肿瘤或引流淋巴结,许多细胞就会抑制和损害其效应器功能。这些细胞包括调节性T细胞、髓源性抑制细胞、II型肿瘤相关巨噬细胞、肿瘤基质细胞(例如成纤维细胞和内皮细胞)和许多其他细胞。此外,一些免疫调节细胞因子,如IL-10、TGF-β和血管内皮生长因子(VEGF),它们直接抑制T细胞的激活、效应器功能和增殖。除了细胞因子外,TME中还释放了许多其他可溶性免疫抑制介质,例如吲哚胺2,3-二加氧酶(IDO)及其代谢产物犬尿氨酸或肿瘤释放的肿瘤代谢产物。
最后,除了TME可以对T细胞施加直接免疫介导的抑制外,纤维化和血管化不良还可以在肿瘤内形成一个不利于最佳T细胞活化或增殖的物理环境。这些物理条件都会损害新抗原特异性T细胞的活性,这突出表明需要新的方法来调控肿瘤微环境以获得更有效的T细胞反应。
肿瘤可及性
在诱导强大的新抗原特异性T细胞反应后最关键的步骤是肿瘤的可及性。换句话说,克服这些T细胞被排除在“冷”肿瘤之外是一个重大挑战。这种排斥可能是由于多种机制造成的,包括物理屏障、血管形成不良以及缺乏T细胞化学吸引。
癌症中物理屏障的一个典型的例子是胰腺导管腺癌,其表现为纤维增生,其特征是广泛的纤维化和细胞外基质(ECM)沉积。重要的是,T细胞在肿瘤内的迁移和靶点识别在这种肿瘤中受到物理限制。克服这一问题的策略可能包括阻断信号通路,如TGF-β、骨形态发生蛋白、Wnt-β-连环蛋白以及Hippo和粘着斑激酶信号通路。在一些癌症中还观察到血管形成不良,如胰腺导管腺癌,甚至会影响药物输送
T细胞耗竭和功能障碍
最后,当治疗性新抗原疫苗引发抗肿瘤T细胞免疫时,这些T细胞在面对高肿瘤负荷时可能会功能失调并耗竭。T细胞耗竭的特征是效应器功能丧失、增殖能力降低、抑制性受体表达增加和细胞死亡倾向,这种耗竭是由持续的抗原和慢性TCR刺激引起的。当疫苗诱导的T细胞进入肿瘤部位时,预计也会遇到类似的情况。如果没有任何补救措施,这种T细胞耗竭将限制新抗原疫苗的益处。将T细胞诱导与ICB相结合的治疗方案可能是解决这一问题的一部分。
晚期和转移性癌症
仍然存在的一个问题是,新抗原疫苗是否对晚期疾病(如大肿瘤或转移性癌症)有效。理想情况下,新抗原疫苗用在转移性疾病之前可切除的小肿瘤,这很可能为患者提供最大的益处,因为肿瘤载量低,而免疫力高,可以防止复发。
在大肿瘤的情况下,许多已经提到的疫苗效力障碍将被放大。此外,不同的器官微环境和转移性TME的差异可能会给疫苗对抗转移的效力增加额外的障碍。在这些情况下,联合疗法可能具有特别的价值。在晚期或转移的情况下,标准护理治疗、免疫疗法和疫苗接种之间的这种强有力的联合治疗很可能是非常必要的。
小结
总之,在开发发现新抗原并引发有效的新抗原特异性抗肿瘤T细胞免疫的技术方面已经取得了相当大的进展,但疫苗要发挥抗癌功效,仍有许多障碍需要克服。肿瘤疫苗接种和癌症免疫学领域需要为这些挑战提供创新的解决方案,以使新抗原疫苗展示其真正潜力,并为癌症患者提供临床益处。
参考文献
1.Challenges in developing personalized neoantigen cancer vaccines. Nat Rev Immunol.2023 Oct 2.
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言



