虎克之后350年,科学家居然发现“细胞蛇”?
2016-06-28 刘冀珑(牛津大学MRC Programme Leader,上海科技大学任教授) 知识分子

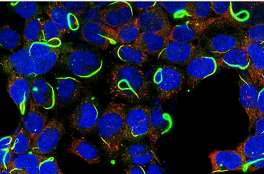
►人胚肾细胞里的CTPS(绿色)和IMPDH(红色)两种细胞蛇 (Chang et al., 2015)。细胞长成一张脱掉英国的欧洲地图 ● ● ● 1生命是什么? ►《时间表:世界历史》。刘冀珑 摄 在我的办公桌上放着从牛津慈善商店淘的一本二手书Time line: History of the World,书名的中文意思是“时间表:世界历史”。在
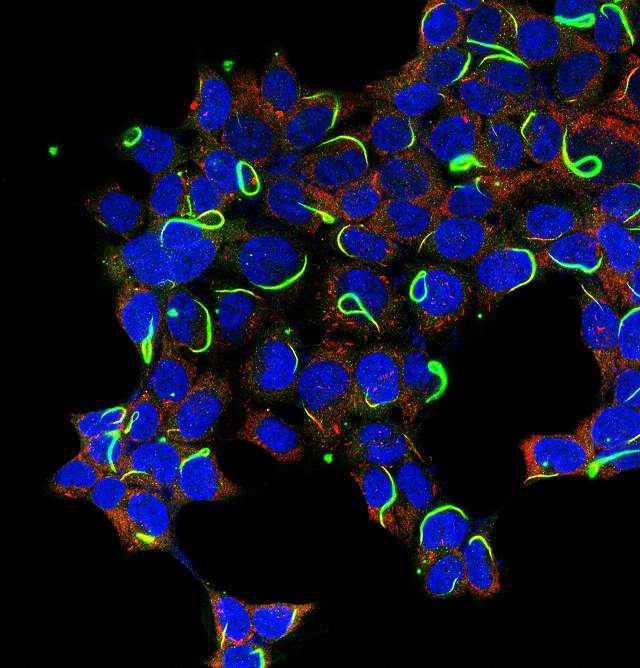
►人胚肾细胞里的CTPS(绿色)和IMPDH(红色)两种细胞蛇 (Chang et al., 2015)。细胞长成一张脱掉英国的欧洲地图
● ● ●
1生命是什么?

►《时间表:世界历史》。刘冀珑 摄
在我的办公桌上放着从牛津慈善商店淘的一本二手书Time line: History of the World,书名的中文意思是“时间表:世界历史”。在喝咖啡的时候,我很喜欢翻阅这本书。作者把世界历史以时间表的形式简单列出,看上去惊天动地的事在历史的沉淀下也不过寥寥数笔。该书的第一条就是公元前137亿年宇宙始于大爆炸。
大约46亿年前,太阳地球月亮相继诞生。
大约38亿年前,第一个细胞在地球上生成,标志着生命的起源。
1943年2月,理论物理学家薛定谔在爱尔兰都柏林的三一学院做了一场报告,主题是:“在活的生物的有限的空间里发生的时空上的事件如何藉由物理和化学来解释”。
主题够拗口的,但没吓住来捧场的400个听众。薛定谔在他的演讲中讲到,生命的一个重要特征是秩序。一年后,他根据这场报告整理出版了一本小书,书名叫What Is Life?(《生命是什么?》)。
这本书吹响了用物理方法研究生命科学的号角。有许多年轻的物理学家受此影响转而研究生物学,其中包括剑桥大学的克里克(Francis Crick)。在35岁“高龄”攻读生物学博士学位期间,克里克与一位美国来的23岁的博士后沃森(James Watson)相遇。他们在剑桥的老鹰酒吧常常一起饮酒聊天。1953年他们揭示了DNA双螺旋结构,标志着分子生物学的纪元。
2细胞是什么?
1665年,牛津大学校友虎克(Robert Hooke)出版了一本书Micrographia(《显微图谱》)。虎克绘画功力了得,同时他折腾出了一个“玩具”——显微镜。虎克拿着他的显微镜,看见什么小东西就拿来看看,然后画下来,并用文字详尽描述。虎克观察的东西千奇百怪。比如,他观察过针尖,随后写一页纸描述针尖在显微镜下的样子。
他还细致观察了厕所小便池里的结晶,跳蚤和虱子等。有一次,他喝完葡萄酒,对着软木塞就是一顿细看。他发现软木塞有很多小室,有些象监狱的牢房,或是修道院的卧室。于是他称这些结构为“Cell” ,英文里是小房间的意思,中文翻译成“细胞”。
后来别的科学家发现植物,动物的组织里都有这些小室,于是提出细胞是生命的基本单位。细胞学说正式确立。
3细胞器是什么?
如薛定谔所言,与非生命相比,生命增加了秩序和复杂性。对于细胞来说,增加秩序和复杂性的一个方式是区域化,就是不同的生物过程和生物分子在细胞里有其独特的定位和结构。
或许我们可以把细胞比喻成一所学校。我们知道,学校里有教室,办公室,实验室,操场和宿舍等等功能结构不同的区域。自从虎克发现细胞以来,尤其是在过去200年里,科学家在细胞里发现了不同的区域。我们把细胞里不同的区域称为细胞器,就好像是细胞里功能不同的器官。
地球早期出现的细胞没有细胞核,我们称之为原核细胞。后来进化的过程中,有一些细胞出现细胞核,这些细胞我们称之为真核细胞。原核生物和真核生物大概在30亿年之前就在进化树上分开了。
1897年意大利科学家高尔基在细胞里用银染法发现了多层网状结构。次年这个结构被称为高尔基体。在此后的50年里,学术界一直有人怀疑高尔基体是否真实存在,认为高尔基体是细胞受固定液影响的假象。
直到1950年代电子显微镜的应用,很多细胞学家在不同的细胞里发现类似的结构,高尔基体是真实的细胞器才获得普遍承认。在过去一百多年中,对于高尔基体研究让我们知道这是一个非常重要的细胞器 。
另一个细胞器经历颇为曲折。纤毛,100多年前被发现。后来很多细胞里都发现有纤毛。但是绝大多数人认为纤毛没有什么用。因为大家一支找不到纤毛的确切功能,很多研究纤毛的科学家没有饭吃,转行做其他研究。在低潮的时候,很少的几个实验室仍然坚持做下去。
最近二十年有了转机。现在大家知道身体中几乎每个细胞都有纤毛。纤毛是信号转导的中心。发育期间纤毛的转动决定了我们身体内脏的左右不对称。纤毛功能失常会导致疾病。纤毛成为细胞生物学的一个热门方向。
线粒体从被发现迄今都是细胞生物学领域的明星,常青树,不倒翁。线粒体被称为细胞的能量工厂。生产作为能量载体的一种核苷酸ATP(三磷酸腺苷),是线粒体的一个重要功能。
你的每一个动作都会消耗ATP。你从饮食中获得营养。营养会通过线粒体里的ATP合成酶生产ATP。ATP除了作为很多代谢反应的能量载体之外,也是合成核酸RNA和DNA的重要零件。合成RNA的四大基本核苷酸分别是ATP, CTP(三磷酸胞苷),UTP(三磷酸尿苷)和GTP(三磷酸鸟苷)。
在动物细胞,ATP合成酶主要定位在线粒体里。在植物细胞,ATP合成酶除在线粒体里之外,也定位在叶绿体里。近60年以来对于合成其他三种核苷酸起关键作用的代谢酶也获得很深入的生物化学研究。
CTP合成酶(CTPS)以UTP为底物生产CTP。CTP合成的关键酶是由肌苷一磷酸脱氢酶(IMPDH)。尽管对CTPS和IMPDH的生物化学研究十分清晰,过去几十年以来我们对于它们在细胞中的定位并不清楚。
4细胞蛇是蛇吗?
过去几年里我做了几十场学术报告,很多是关于细胞蛇的报告。星期天早晨在电梯里偶遇一个研究生。他问我,“老师,您是研究蛇的吗?”
2010年的夏天,三组科学家独立报道了CTPS在果蝇,细菌和芽殖酵母中能形成纤维状结构 。一年后, CTPS的这种形成纤维的特点在人类细胞中得到证实。
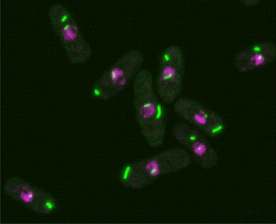

►果蝇细胞里的细胞蛇(绿色),Liu 2010
这种纤维状结构因为其形状和蛇类似,我称之为Cytoophidium。这个词是希腊语“cyto” (细胞)和“ophidium”(蛇)两个词组合起来的,所以中间有两个“o”。“Cytoophidium”是单数,它的复数形式是“Cytoophidia”。细心一点的读者可能会发现,“Cytoophidium”这个单词中可以找到C,T,P三个字母。
这是巧合吗?
现在可以回答电梯口那位同学了,“同学,我不是研究蛇的,我是研究细胞蛇的”。细胞蛇不是蛇,它是细胞里形态类似蛇,含有CTPS或其他代谢酶的一种结构。
5细胞蛇是怎样被发现的?
2007年8月,我从美国跨过大西洋来到英国,在牛津大学组建实验室,做博士生导师。因为某种原因,我接手了转来的三个博士生。其中一位学生还有一年应该毕业。我感觉学生的实验数据仍然不够,觉得应该帮助她。于是我亲手做实验,希望加快学生课题的进度。
我们对果蝇生殖细胞里的两种球形结构比较有兴趣,想知道二者功能上的关系。这两种球形结构分别称为U body (U小体)和P body (P小体)。U是球形。P近乎球形,有时不很规则。
我们最初的努力集中于一个翻译起始复合物的结合蛋白,该蛋白的名字叫Cup(杯子)。因为如果该基因突变了,果蝇的卵泡会变形,象杯子一样。
Cup蛋白一般用作P小体的标记物。我使用来自不同来源的4个抗Cup 蛋白的抗体做免疫染色。其中3个如我预期,染出P小体。但是第四个抗体除了染出P小体之外,还染出了一种神秘的丝状结构。这种结构在果蝇的护理细胞,卵母细胞和卵泡细胞都有。
这是一个很意外的观察结果。
由于不想让学生的课题担当风险,我决定亲自跟从这个偶然观察。我推测这些丝状结构是纤毛。接下来两年里,我做了不同实验试图证实这个推测。但是证据一直不能让人信服。
到了2009年,我的推测被实验证实是错的。这些结构不是纤毛。既然不是纤毛,我觉得有必要给它们取个名字。因为它们的丝状蛇形形式,我将这些新颖结构称作Cytoophidium(细胞蛇)。
我所观察的细胞蛇与两株绿色荧光蛋白(GFP)标记的CTPS蛋白的分布很像 。经过实验,我发现标记细胞蛇的抗体和标记CTPS的GFP定位吻合。
为了确保万无一失,我用三个特异性地识别CTPS蛋白不同区域的抗体进行免疫染色。所有结果无一例外,定位吻合。至此,我很肯定所观察到的细胞蛇里含有CTPS。
我以为细胞蛇是生殖细胞特有的结构,但在用果蝇其他组织细胞进行的实验结果显示,含有CTPS的细胞蛇存在于许多组织,包括脑,肠,气管,睾丸,附腺,唾液腺和淋巴腺。此外,在除果蝇外的其他物种细胞中也可观察到细胞蛇。
2010年5月,我发表了一篇报道果蝇细胞里的这些观察结果的文章。这是“Cytoophidium”(细胞蛇)这个词第一次出现在公共知识区域。
两个月后,CTPS被描述在细菌中形成丝状结构。又过了一个月,发表于2010年8月的一篇论文表明,在芽殖酵母蛋白质CTPS也能形成丝状结构。
在过去的六十年,科学家已经对CTPS蛋白的生物化学经过了深入的研究,因而在2010年夏天独立发表的这三篇论文对研究CTPS的科学家来讲既是出人意外,又是非同寻常的——CTPS分子可以形成细胞蛇这样一个丝状结构。
随后,我的实验室和另一个实验室又分别证实CTPS也可以在人类细胞形成细胞蛇。我们进而发现裂殖酵母里细胞蛇的存在。在人、小鼠和酵母菌细胞里,细胞质和细胞核里我们均发现有细胞蛇。
►裂殖酵母中的细胞蛇(绿色为CTPS,紫色为细胞核)(zhang et al., 2014)。
因此,细胞蛇代表了一种新发现的,在原核细胞和真核细胞里普遍存在的,进化上高度保守的细胞结构。
6细胞蛇是细胞器吗?
按字面意思,细胞器是指“细胞的器官”。在细胞里的独特而且广泛存在的结构,应该都属于细胞器的范畴。
定义一个结构为细胞蛇必须满足三个标准。 首先,它是丝状结构(相对于球形结构如P小体,U小体)。第二,该结构通常含有代谢酶(有别于经典细胞骨架的微管,微丝和中间丝)。第三,该结构缺乏膜(不同于有膜的细胞器如线粒体,内质网,高尔基体和纤毛)。
这个定义满足了定义细胞蛇为细胞器独特性方面的要求。之前我们说过, 细胞蛇在原核细胞和真核细胞里普遍存在的,进化上高度保守。
历史上由于电子显微镜下容易辨别有膜的细胞结构,人们过去常常认为细胞器必须有膜包裹。比如线粒体,细胞核,纤毛,溶酶体等。
随着细胞生物学技术的提高,细胞内没有膜的结构逐渐被发现。比如我们之前感兴趣的在细胞质的U小体和P小体,还有细胞核里的柯哈小体(Cajal body)和组蛋白基因位点小体(histone locus body),都是无膜的结构。细胞蛇也没有膜包裹。一般来讲,我们认为这些结构是新类型的无膜细胞器,与传统意义的被膜包裹的细胞器不同。按照这个定义,细胞蛇是一种无膜细胞器。
7为什么会有细胞蛇?
坦率地讲,我不知道答案。我们正在探索这个问题。几个特点需要考虑。首先,细胞蛇的形成是一个非常古老的现象。细菌和人类细胞的祖先在进化树上30亿年前就已经分离,但他们的CTPS分子都能形成细胞蛇。
其次,细胞蛇广泛存在于不同细胞里。因此,细胞蛇可能参与到细胞组织功能的一些基本机制中。
第三,细胞蛇在原核与真核细胞中的普遍存在表明, CTPS形成的细胞蛇并没有在进化的压力下被抛弃。这表明细胞蛇对于一个生物体的繁殖和存活是有益的。
8为什么要研究细胞蛇?
含CTPS纤维状结构在不同物种中普遍存在表明,细胞蛇的形成可能具有极其重要的生物学功能,并可能参与到细胞生产CTP和其他核苷酸的共同管理策略中 。极为重要的是,与经典的细胞器类似,细胞蛇的调控失常会与人类疾病有关联。
2014年几个研究组对细胞蛇或类似的结构对于代谢调控的作用和机制做出了详细研究。这四个研究均认为CTPS聚合形成蛇形结构可以快速让酶活性降低。最近的研究也表明,细胞蛇是动态的结构,与代谢状态和环境条件有关 (Shen et al., 2016)。
GTP合成的限速反应由IMPDH催化。有趣的是,IMPDH形成丝状结构,非常类似含CTPS的细胞蛇。进一步研究表明这两种细胞蛇相互独立又互相作用。
最近的研究表明,越来越多的代谢酶可以在特定条件下形成细胞蛇或同类结构。对4159个芽殖酵母蛋白进行筛选,我们找到至少23个蛋白可以形成类似于细胞蛇的纤维状结构,与之前的一个稍小规模的研究吻合。
这些具有形成纤维状结构的蛋白大多是代谢酶,它们集中在与翻译起始,葡萄糖和氮代谢相关的几个通路上。
2014年几个研究组对细胞蛇或类似的结构对于代谢调控的作用和机制做出了详细研究。这四个研究均认为CTPS聚合形成蛇形结构可以快速让酶活性降低。最近的研究也表明,细胞蛇是动态的结构,与代谢状态和环境条件有关 (Shen et al., 2016)。
GTP合成的限速反应由IMPDH催化。有趣的是,IMPDH形成丝状结构,非常类似含CTPS的细胞蛇。进一步研究表明这两种细胞蛇相互独立又互相作用。
最近的研究表明,越来越多的代谢酶可以在特定条件下形成细胞蛇或同类结构。对4159个芽殖酵母蛋白进行筛选,我们找到至少23个蛋白可以形成类似于细胞蛇的纤维状结构,与之前的一个稍小规模的研究吻合。
这些具有形成纤维状结构的蛋白大多是代谢酶,它们集中在与翻译起始,葡萄糖和氮代谢相关的几个通路上。

►遗传学报(JGG)2016年6月封面,显示芽殖酵母中“细胞蛇结构”蛋白之间的相互关系
1978年,威廉姆斯和他的同事发现,在肝癌中CTPS活性升高。随后的研究表明,CTP水平失控和增加CTPS活性是许多种癌症(例如白血病,肝细胞瘤和结肠癌)的一个特征。我们正试图探索细胞蛇组装和癌症发生的关系。CTPS1对淋巴细胞增殖非常关键。此外,CTPS对于生殖和大脑发育很重要。
最近我们在果蝇中的研究显示,原癌基因Myc和CTPS相互影响,一方面Myc调控CTPS组装成细胞蛇,另一方面CTPS影响Myc介导的对细胞大小的控制。CTPS组装成细胞蛇也受非受体酪氨酸激酶Ack和E3泛素连接酶Cbl的调控。
CTPS也一直是药物开发中针对病毒性疾病和寄生虫病(例如非洲昏睡病,疟疾和感染性失明)的有吸引力的靶标。 CTPS组装成细胞蛇的特性或许可以考虑应用于治疗病毒感染或抗寄生虫靶向药物的设计。
细胞蛇是细胞生物学的新领域。多学科手段和尖端技术将加速我们对细胞蛇的系统理解。对细胞蛇的研究尚处于起步阶段。许多悬而未决同时又意义重大的问题,预示着细胞蛇的研究将成为一件非常令人兴奋的事情。
作者简介
刘冀珑,1971年生于江西九江。细胞蛇(Cytoophidia)的发现者。牛津大学MRC Programme Leader。2016年6月入职上海科技大学任教授。2007年8月,刘冀珑入职英国牛津大学医学部的生理、解剖与遗传系,成为博士生导师,以果蝇为模式动物专注于细胞蛇的生物学研究。刘冀珑早年毕业于北京林业大学(本科),中国农业大学(硕),中国科学院动物研究所(博)。他博士后合作导师依次是康涅狄格大学的胚胎学家杨向中教授和卡内基研究所细胞生物学家Joseph Gall教授。
原始出处:
1.AUGHEY, G. N., GRICE, S. J. & LIU, J. L. 2016. The interplay between Myc and CTP synthase in Drosophila. PLoS Genet, 12, e1005867.
2.AUGHEY, G. N., GRICE, S. J., SHEN, Q. J., XU, Y., CHANG, C. C., AZZAM, G., WANG, P. Y., FREEMAN-MILLS, L., PAI, L. M., SUNG, L. Y., YAN, J. & LIU, J. L. 2014. Nucleotide synthesis is regulated by cytoophidium formation during neurodevelopment and adaptive metabolism. Biol Open, 3, 1045-56.
3.AUGHEY, G. N. & LIU, J. L. 2016. Metabolic regulation via enzyme filamentation. Crit Rev Biochem Mol Biol, 51, 282-93.
4.AZZAM, G. & LIU, J. L. 2013. Only one isoform of Drosophila melanogaster CTP synthase forms the cytoophidium. PLoS Genet, 9, e1003256.
5.BARRY, R. M., BITBOL, A. F., LORESTANI, A., CHARLES, E. J., HABRIAN, C. H., HANSEN, J. M., LI, H. J., BALDWIN, E. P., WINGREEN, N. S., KOLLMAN, J. M. & GITAI, Z. 2014. Large-scale filament formation inhibits the activity of CTP synthetase. Elife, 3, e03638.
6.BUSZCZAK, M., PATERNO, S., LIGHTHOUSE, D., BACHMAN, J., PLANCK, J., OWEN, S., SKORA, A. D., NYSTUL, T. G., OHLSTEIN, B., ALLEN, A., WILHELM, J. E., MURPHY, T. D., LEVIS, R. W., MATUNIS, E., SRIVALI, N., HOSKINS, R. A. & SPRADLING, A. C. 2007. The carnegie protein trap library: a versatile tool for Drosophila developmental studies. Genetics, 175, 1505-31.
7.CARCAMO, W. C., SATOH, M., KASAHARA, H., TERADA, N., HAMAZAKI, T., CHAN, J. Y., YAO, B., TAMAYO, S., COVINI, G., VON MUHLEN, C. A. & CHAN, E. K. 2011. Induction of cytoplasmic rods and rings structures by inhibition of the CTP and GTP synthetic pathway in mammalian cells. PLoS ONE, 6, e29690.
8.CHANG, C. C., LIN, W. C., PAI, L. M., LEE, H. S., WU, S. C., DING, S. T., LIU, J. L. & SUNG, L. Y. 2015. Cytoophidium assembly reflects upregulation of IMPDH activity. J Cell Sci, 128, 3550-5.
9.CHEN, K., ZHANG, J., TASTAN, O. Y., DEUSSEN, Z. A., SISWICK, M. Y. & LIU, J. L. 2011. Glutamine analogs promote cytoophidium assembly in human and Drosophila cells. J Genet Genomics, 38, 391-402.
10.GOU, K. M., CHANG, C. C., SHEN, Q. J., SUNG, L. Y. & LIU, J. L. 2014. CTP synthase forms cytoophidia in the cytoplasm and nucleus. Exp Cell Res, 323, 242-53.
11.HOOKE, R. 1665. Micrographia: Or Some Physiological Descriptions of Minute Bodies Made by Magnifying Glasses. Jo. Martyn & Ja. Allestry, London.
12.INGERSON-MAHAR, M., BRIEGEL, A., WERNER, J. N., JENSEN, G. J. & GITAI, Z. 2010. The metabolic enzyme CTP synthase forms cytoskeletal filaments. Nat Cell Biol, 12, 739-46.
13.KEPPEKE, G. D., CALISE, S. J., CHAN, E. K. & ANDRADE, L. E. 2015. Assembly of IMPDH2-based, CTPS-based, and mixed rod/ring structures is dependent on cell type and conditions of induction. J Genet Genomics, 42, 287-99.
14.KERR, G. 2007. Time line: History of the World. Futura, London pp1-157.
15.LEE, L., DAVIES, S. E. & LIU, J. L. 2009. The spinal muscular atrophy protein SMN affects Drosophila germline nuclear organization through the U body-P body pathway. Dev Biol, 332, 142-55.
16.LIU, J. L. 2010. Intracellular compartmentation of CTP synthase in Drosophila. J Genet Genomics, 37, 281-96.
17.LIU, J. L. 2011. The enigmatic cytoophidium: compartmentation of CTP synthase via filament formation. Bioessays, 33, 159-64.
18.LIU, J. L. 2016. The cytoophidium and its kind: Filamentation and compartmentation of metabolic enzymes. Annu Rev Cell Dev Biol, 32, (in press).
19.LIU, J. L. & GALL, J. G. 2007. U bodies are cytoplasmic structures that contain uridine-rich small nuclear ribonucleoproteins and associate with P bodies. Proc Natl Acad Sci U S A, 104, 11655-9.
20.LIU, J. L., MURPHY, C., BUSZCZAK, M., CLATTERBUCK, S., GOODMAN, R. & GALL, J. G. 2006b. The Drosophila melanogaster Cajal body. J Cell Biol, 172, 875-84.
21.NOREE, C., MONFORT, E., SHIAU, A. K. & WILHELM, J. E. 2014. Common regulatory control of CTP synthase enzyme activity and filament formation. Mol Biol Cell, 25, 2282-90.
22.NOREE, C., SATO, B. K., BROYER, R. M. & WILHELM, J. E. 2010. Identification of novel filament-forming proteins in Saccharomyces cerevisiae and Drosophila melanogaster. J Cell Biol, 190, 541-51.
23.PETROVSKA, I., NUSKE, E., MUNDER, M. C., KULASEGARAN, G., MALINOVSKA, L., KROSCHWALD, S., RICHTER, D., FAHMY, K., GIBSON, K., VERBAVATZ, J. M. & ALBERTI, S. 2014. Filament formation by metabolic enzymes is a specific adaptation to an advanced state of cellular starvation. Elife, 3, e02409.
24.Schrödinger, E. 1944. What Is life? Cambridge University Press, Cambridge.
25.Shen, Q. J., Kassim, H., Huang, Y., Li, H., Zhang, J., Li, G., Wang, P. Y., Yan, J., Ye, F. F. & Liu, J. L. 2016. Filamentation of metabolic enzymes in Saccharomyces cerevisiae. J Genet Genomics 43, 393-404.
26.STROCHLIC, T. I., STAVRIDES, K. P., THOMAS, S. V., NICOLAS, E., O'REILLY, A. M. & PETERSON, J. R. 2014. Ack kinase regulates CTP synthase filaments during Drosophila oogenesis. EMBO Rep, 15, 1184-91.
27.TASTAN, O. Y. & LIU, J. L. 2015. CTP Synthase Is Required for Optic Lobe Homeostasis in Drosophila. J Genet Genomics, 42, 261-74.
28.WANG, P. Y., LIN, W. C., TSAI, Y. C., CHENG, M. L., LIN, Y. H., TSENG, S. H., CHAKRABORTY, A. & PAI, L. M. 2015. Regulation of CTP Synthase Filament Formation During DNA Endoreplication in Drosophila. Genetics, 201, 1511-23.
29.ZHANG, J., HULME, L. & LIU, J. L. 2014. Asymmetric inheritance of cytoophidia in Schizosaccharomyces pombe. Biol Open, 3, 1092-7.
版权声明:
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言




中国学者顶起
153
也是原创发现
178