
Toll样受体的辅助分子及其功能
2024-12-06 小药说药 小药说药 发表于陕西省

本文介绍模式识别受体中 Toll 样受体相关内容,阐述细胞表面 TLR、内体 TLR 的各类辅因子,以及 TLR 的伴侣、转运和加工因子的作用机制,强调其对理解 TLR 途径及潜在疗法设计的重要性。
模式识别受体(PRRs)是一类先天免疫受体,是病原体相关分子模式(PAMP)的传感器;同时PRRs还可以识别响应压力或组织损伤而释放的内源性分子,从而充当损伤相关分子模式(DAMP)的传感器。Toll样受体(TLR)是识别许多病原体衍生大分子的PRR,TLR激活导致转录因子核因子-κB(NF-κB)和干扰素调节因子(IRFs)的激活,分别产生促炎细胞因子和I型干扰素(IFNs)。
TLRs是I型跨膜蛋白,由胞外结构域中的富含亮氨酸重复序列(LRR)、单个跨膜结构域和参与信号衔接分子募集的细胞质Toll/IL-1受体(TIR)结构域组成。人类表达10个功能性TLR(TLR1至TLR10),TLR1、TLR2、TLR4、TLR5、TLR6位于细胞膜上,在那里它们识别病原体表面的分子成分。TLR3、TLR7、TLR8和TLR9存在于细胞内内涵体膜上,在那里它们介导核酸的识别。
TLR形成异二聚体或同源二聚体作为触发信号的手段,细胞外和内体TLR的外结构域序列都是同源的,这一特征与它们识别的配体的多样性形成鲜明对比。配体区分的一种模式依赖于TLR外结构域中存在残基的差异,然而氨基酸变异和异二聚体的形成只能为识别不同的TLR配体提供有限的支持。因此,必然存在另一种反映TLR配体组成复杂性和多样性的机制,以确保正确的PAMP检测和自我/非自我区分。
特定的辅助蛋白或辅因子可以发挥这一作用:它们是TLR功能所必需的,它们与TLR或TLR配体相互作用,辅助TLR的激活。由于它们在TLR功能中的重要作用,靶向这些辅助蛋白可能有利于旨在操纵TLR激活用于治疗应用的策略。
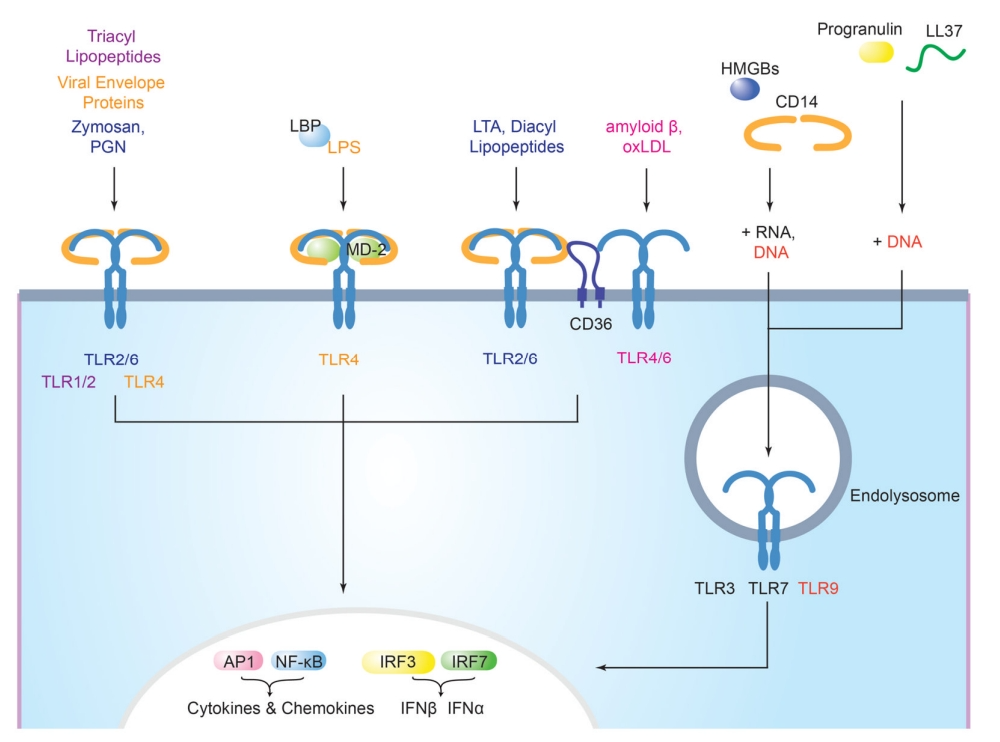
LBP
LPS结合蛋白(LBP)是一种481个氨基酸的糖蛋白,与来源于革兰氏阴性菌外膜的脂多糖(LPS)具有高亲和力。在细胞对LPS的应答过程中,LBP主要起催化作用,即LBP将LPS单体从其多聚体中转运到CD14,加速LPS与 CD14的结合,1个分子LBP可促使上百分子LPS与CD14结合,介导TLR4对LPS的反应性。
此外,LBP还可以与脂磷壁酸(LTA)、肽聚糖和脂肽结合并将其转移到CD14,这表明LBP不仅可以帮助TLR4的功能,还可以帮助TLR1、TLR2和TLR6的功能。因此,LBP介导了对来源于革兰氏阴性菌和革兰氏阳性菌的PAMP的先天免疫反应。
MD2
MD2是一种160个氨基酸的糖基化可溶性蛋白,与TLR4的细胞外结构域结合,它是体内TLR4依赖性LPS反应所必需的。TLR4-MD2的晶体结构显示了MD2如何促进TLR4的功能:LPS将其六条脂质链中的五条埋入MD2的疏水口袋中,两个MD2-LPS复合物对于桥接两个TLR4分子至关重要。由于TLR4-MD2异二聚体中的两个TLR4分子具有有限的直接相互作用,MD-2对于TLR4的配体结合和二聚化都是必不可少的。
CD36
CD36是在脂筏中发现的B型清道夫受体家族的双跨膜糖蛋白。CD36与TLR2-TLR6异二聚体的功能有关,CD36增强了对某些TLR2-TLR6配体的免疫反应。在体内,CD36的缺乏会导致革兰氏阳性金黄色葡萄球菌感染的易感性增加。此外,CD36还通过TLR4-TLR6异二聚体的组装介导对氧化低密度脂蛋白(oxLDL)和淀粉样β纤维的炎症反应。
CD36如何介导TLR2-TLR6和TLR4-TLR6的功能尚不完全清楚,但CD36的C末端似乎起着重要作用。酪氨酸激酶LYN与CD36之间的相互作用需要CD36的残基460-463,抑制LYN激酶活性会损害CD36与TLR4-TLR6的结合,并阻断NF-κB对oxLDL的激活。因此,将LYN募集到CD36的C末端对于形成功能性TLR4-TLR6-CD36信号复合物非常重要。
CD14
CD14是一种375个氨基酸的富含亮氨酸的糖蛋白,以可溶形式存在于血液中,或作为髓系细胞上的糖基磷脂酰肌醇(GPI)锚定膜蛋白存在。CD14与多种TLR配体相互作用,增强其激活TLR的能力。重组CD14的直接结合研究表明,CD14具有结合多种微生物产物的不同寻常的能力,包括LPS、肽聚糖、Pam3CSK4、polyI:C和CpG DNA。
CD14首先与TLR4介导的免疫反应有关,作为对LPS的反应,CD14是TRIF依赖性信号传导所必需的,在低剂量下是MyD88依赖性信号转导所必需的。CD14还增强了对内体TLR配体polyI:C、咪喹莫特和CpG DNA的免疫反应,CD14可能促进核酸的普遍内化。
CD14除了结合多种配体的能力,还介导了TNF的产生,以响应TLR2-TLR6配体MALP2、LTA、酵母聚糖A和Pam2CSK4,并参与了TLR介导的对各种病毒的免疫反应,CD14究竟如何参与这些过程仍有待确定,但和LPS和核酸类似,CD14可能介导配体与几种TLR的相互作用。
TRIL
TRIL是一种由811个氨基酸组成的I型跨膜蛋白,在其细胞外结构域中含有12个重复的富含亮氨酸序列。研究表明,TRIL介导TLR3和TLR4信号,但不介导TLR2或TLR9信号。TRIL与LPS、TLR3和TLR4免疫共沉淀,表明TRIL参与配体递送。
Granulin
颗粒蛋白(Granulin)是一种富含半胱氨酸的糖基化多功能蛋白,可以与TLR9相互作用。添加外源性颗粒蛋白前体可增强RAW巨噬细胞对合成寡脱氧核苷酸CpG-B和CpG-C的TNF分泌。在Grn-/-小鼠中,CpG ODN与TLR9的结合受损,因此颗粒蛋白可能有助于CpG递送到溶酶体隔室。这些结果表明,颗粒蛋白有助于CpG的递送,以促进TLR9反应。目前尚不清楚颗粒蛋白是否与表面受体相互作用,以及是什么决定了TLR9对CpG-B和CpG-C而不是CpG-a的反应增强。未来的研究应澄清这些问题,并确定TLR9激活需要哪种形式的颗粒蛋白。
HMGB1
高迁移率族蛋白B1(HMGB1)家族的成员是与染色质相关的核蛋白,参与DNA的转录调控。HMGB1是该家族中研究最多的成员,通过与其受体TLR2、TLR4和晚期糖基化终末产物受体(RAGE)的相互作用介导促炎功能。
HMGB蛋白可能是核酸先天免疫反应的普遍介质。HMGB1结合DNA和RNA,以响应TLR3、TLR7和TLR9的激活,是I型IFN和促炎细胞因子产生所必需的。HMGB1首先被描述为TLR9辅因子,因为它能够结合CpG DNA,与TLR9相互作用,并增强TLR9对CpG的内体递送。外源性添加HMGB1可增强DC和巨噬细胞对CpG的IFN-α和促炎细胞因子的产生。IFN-α的分泌依赖于HMGB1与RAGE的相互作用,RAGE是TLR9的“被动”受体辅因子。HMGB1是TLR9对CpG DNA反应所必需的,由于其结合DNA的能力,可能会加剧自身免疫性疾病。
LL37
LL37是TLR9辅因子,与pDCs中自身DNA向TLR9的递送有关。LL37是一种长37个氨基酸的两亲性肽,在丝氨酸蛋白酶切割其前体后被激活。LL37-DNA复合物对DNA酶的降解具有抗性,并被pDCs内化,定位于早期内体,从那里介导TLR9依赖性IFN-α的产生。目前没有证据表明LL37和TLR9之间存在直接相互作用,这表明LL37在细胞损伤的情况下可能主要作为DNA递送分子。LL37也被证明可以与自身RNA形成复合物,并将这些复合物递送到pDCs,以产生TLR7和TLR8依赖的IFN。LL37在TLR驱动的反应中对宿主防御是否重要仍有待确定。
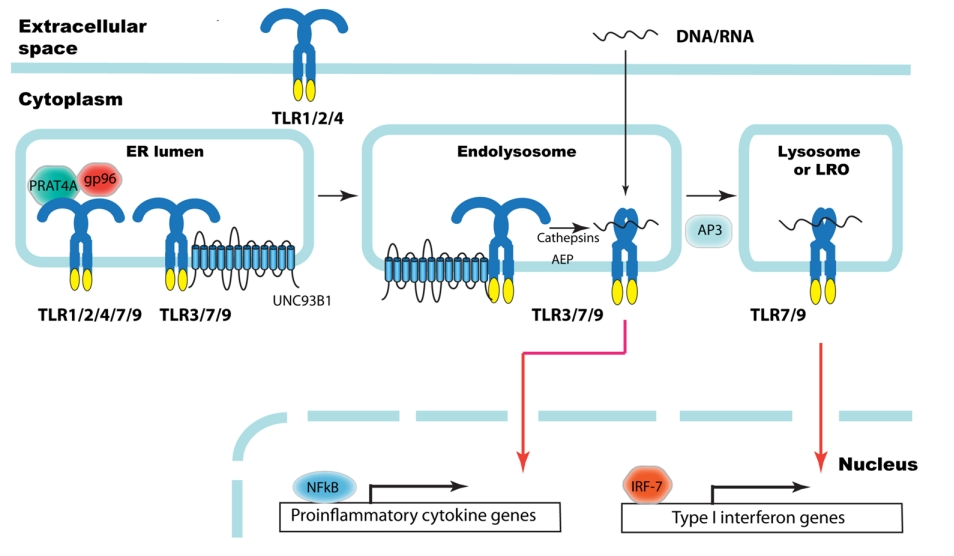
Gp96和PRAT4A
Gp96是介导蛋白质折叠的HSP90家族伴侣的内质网(ER)同源物。Gp96普遍表达,并作为专性可溶性同二聚体存在,每个单元由N端ATP结合结构域、高电荷中间结构域和C端二聚结构域组成。gp96的靶点数量有限,包括整合素、血小板糖蛋白复合物和TLR。TLR1、TLR2、TLR4、TLR5、TLR7和TLR9的功能需要gp96。gp96介导TLR的折叠和成熟,然而gp96究竟在哪个阶段干预TLR折叠,以及如何干预的,尚不清楚。
PRAT4A是一种普遍存在且高度保守的可溶性ER管腔蛋白。与gp96类似,PRAT4A对ER中多种TLR的成熟很重要,PRAT4A和gp96协同工作,以确保TLR的正确折叠。
UNC93B1
UNC93B1是一种ER驻留糖蛋白,是内体TLR反应所必需的。研究表明,UNC93B1与TLR3、TLR7、TLR8、TLR9相互作用,内体TLR的跨膜结构域控制着与UNC93B1的结合。核酸感应TLR必须通过其跨膜结构域与UNC93B1相互作用,以便UNC93B1可以介导其递送到内溶酶体,在那里它们可以结合并响应各自的配体。
AP3
适配蛋白3(AP3)是TLR9运输机制的必需成分。AP家族的成员是四聚体蛋白,它们介导分泌和内吞途径中膜蛋白的选择。AP3由四个亚基组成:δ、μ3A、β3A和σ3,负责将DNA/RNA招募到内体中,以输送到溶酶体和溶酶体相关细胞器(LRO)。
组织蛋白酶
TLR9在到达内溶酶体隔室时进行蛋白水解处理,也可能在具有低pH值和蛋白酶的早期内体中进行。处理TLR的蛋白酶必须与TLR9相互作用,因此被认为是辅因子。组织蛋白酶K缺乏导致BMDC细胞因子对CpG的反应降低,表明组织蛋白酶与TLR9功能有关。组织蛋白酶B、组织蛋白酶L、组织蛋白酶S和组织蛋白酶F被鉴定为与B细胞系TLR9功能相关的因子。此外,单个组织蛋白酶的抑制剂未能完全抑制切割和TLR驱动的反应,表明要获得完整的TLR9活性,需要多种组织蛋白酶的活性。
TLR受体可以识别差异很大的PAMP,但显示出结构保守的外结构域。许多有助于配体区分和受体信号传导的分子已经被鉴定出来,这些分子显示出不同的功能:辅因子、信号适配器以及TLR功能的调节因子。TLR特异性激活的最终结果必须来自这些介质的组合,从而产生复杂的信号。
由于它们对TLR功能的贡献,对有助于激活TLR的辅因子的研究非常重要,有助于我们更好地理解控制先天性和适应性免疫的TLR途径。尽管这些认识是否可以应用于设计新的疗法尚无法衡量,但操纵TLR的其他手段仍然是一个非常理想的目标。
参考文献:
1.Accessory molecules for Toll-like receptors and their function. Nat Rev Immunol.2012 Feb 3;12(3):168-79
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言





#Toll样受体# #辅助分子#
8