肿瘤免疫治疗中检查点反应的原型
2022-09-08 小药说药 小药说药

ICB反应性至少有两种“类型”——I类和II类反应性免疫原型,这些原型的组成部分已经具有明确的预后,这些特征的结合有助于更好地预测ICB的反应性。
前言
目前,癌症患者对免疫检查点阻断(ICB)治疗的反应性是通过不同的个体测量来预测的,其准确度不同,包括肿瘤突变负荷、肿瘤浸润性T细胞密度、树突状细胞频率和检查点配体的表达。这些个体参数是相互关联的,根据细胞类型和基因表达,在ICB反应性患者中形成了两种不同的“反应性”免疫原型。
这些是抗肿瘤免疫的“种子”,并由肿瘤微环境(TME)的特定元素和微生物组的作用支持。尽管消除TME中的“免疫抑制”因素很重要,但了解和分析反应性免疫对于最佳预后以及将其与候选疗法结合以提高肿瘤治愈率至关重要。
免疫原型:功能目标的多重关联状态
在活组织和免疫系统中,细胞状态相互支持。例如,1型常规树突状细胞(cDC1)支持CD8+T细胞效应器功能,而2型常规树突状细胞(cDC2)支持CD4+T细胞。这些细胞类型的功能联系可能代表进化选择的功能配对,以实现特定类型的反应。因此,不同于关注单个细胞类型的状态,应该通过考虑具有连接状态的细胞集合来定义免疫系统及其功能目标,这就是免疫原型。
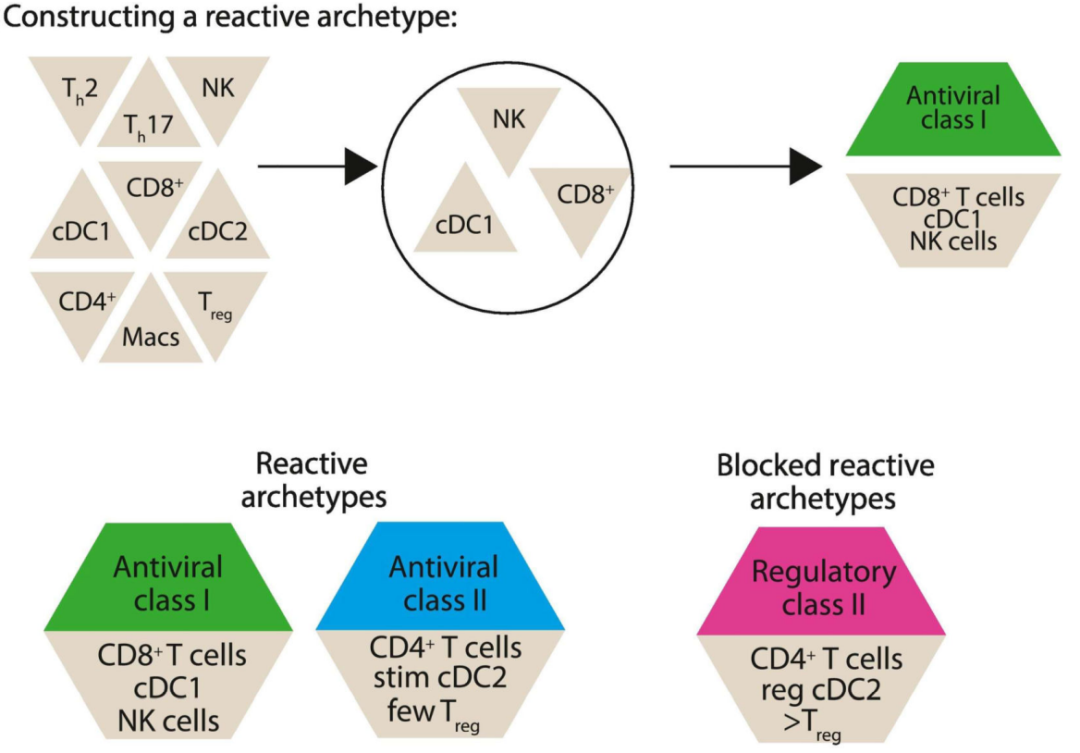
反应性免疫原型是可以介导肿瘤消退的细胞类型的集合,组成反应性免疫原型的细胞数量稀少,可以在主要由显性原型构成的肿瘤中共存。然而,虽然组成反应性原型的细胞在生长中的肿瘤中只有少量存在,但它们是产生免疫反应的种子,ICB最终会增强免疫反应,因此它们的初始存在被认为是至关重要的。
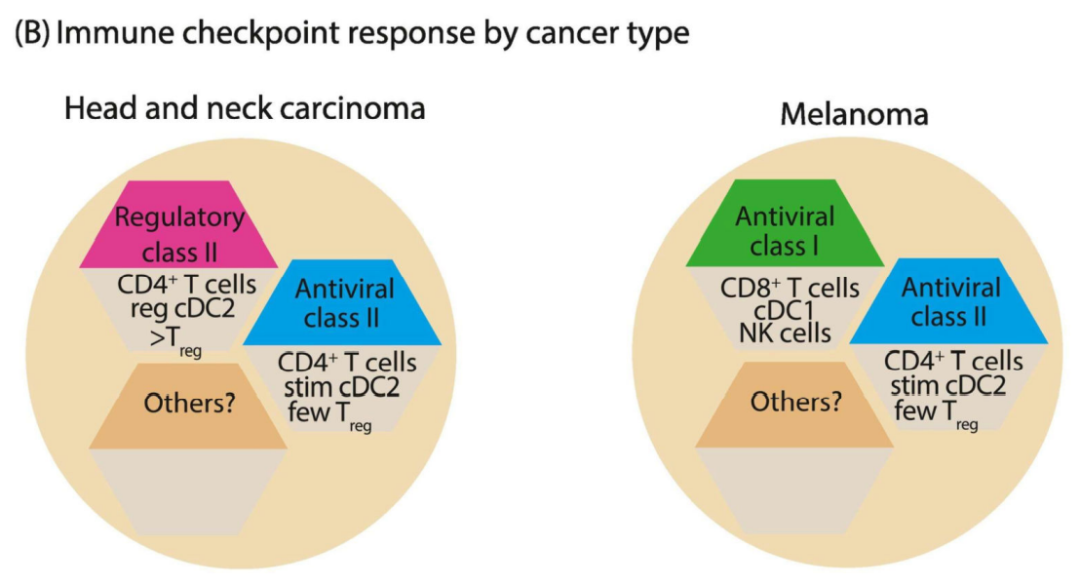
上图概述了对特定癌症类型如何具有支持特定反应原型不同倾向的粗略理解。例如,约40%的黑色素瘤患者呈现I类“反应性”原型,占对PD-1抑制剂有反应的大多数患者,但在头颈部鳞状细胞癌(HNSCC)中未观察到这种情况。然而,在一些黑色素瘤患者中,一种不太常见的反应性原型(II类原型)是由cDC2+/CD4+浸润形成的,这种原型在HNSCC中相对常见。因为这些反应原型是不同的,因此识别这些反应原型将有利于预先确定什么样的ICB治疗方案可能对特定患者最有效。
I类:基于CD8的ICB反应原型
CD8+T细胞的特点是其细胞溶解功能,这是由TCR和肽-MHC I复合物(pMHC)之间的相互作用介导的。CD8+肿瘤浸润淋巴细胞优先识别含有pMHC的自身蛋白,这些蛋白是由肿瘤产生的特异性抗原,区别于自身健康组织。许多肿瘤驻留CD8+T细胞表现出抑制性检查点受体PD-1或CTLA-4的高表达,并具有不同于真正效应或组织驻留记忆T细胞的转录状态。
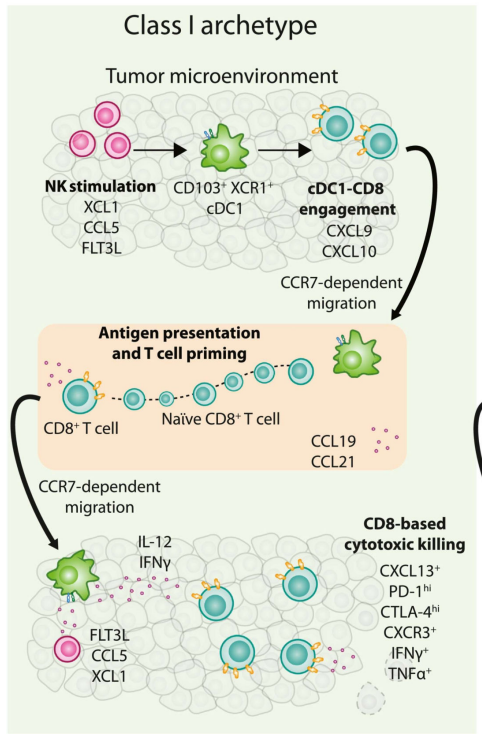
来自小鼠肿瘤和慢性病毒感染模型的证据表明,一些CD8+T细胞亚群的耗竭状态不同,并且有可能在PD-1阻断后重新激活效应器功能。在人类研究中,表达CD103+CD69+的CD8+T细胞亚群,称为组织驻留记忆细胞(TRM),其与多种癌症类型中更好的患者总体生存率相关。与T细胞功能障碍以PD-1高表达为标志的观点相反,PD-1hiTRM细胞数量的增加已被证明可以预测NSCLC患者对ICB的积极反应。这些特征表明,一些CD8+PD-1hiTRM群体,不是一直处于耗竭状态,其可能代表I类反应原型中的关键细胞类型。
这些CD8+TIL是如何激活的?研究发现,小鼠肿瘤中的TME cDC1表达大量刺激性细胞因子IL-12并且在中性pH环境中维持摄入的蛋白质在MHC I上交叉呈现肿瘤抗原的能力,它们对于扩增浸润的T细胞也是必不可少的。从机制上讲, cDC1上调趋化因子受体CCR7,之后cDC1迁移到淋巴结(LN)。cDC1不仅可以直接在LN中激发新的T细胞,还可以通过独特的DC-DC突触传递抗原;然后抗原被转移到CD8+T细胞以及其他DC亚群。这反过来又允许新的CTL的大量激活和增殖。总之,在TME中的cDC1是反应性I类原型的必要组成部分。
最后,第三种细胞类型,自然杀伤(NK)细胞,是I类反应原型的关键组成部分。NK细胞表达大量趋化因子CCL5和XCL1,它们与cDC1上的受体结合,并与肿瘤中的cDC1密切相关。同时,NK细胞分泌Flt3L(cDC1的生长因子),NK细胞频率与人类黑色素瘤中cDC1的数量高度相关。
II类:基于CD4的ICB反应原型
通过过继转移肿瘤反应性CD4+T细胞或选择性地消耗CD4+T细胞,已在多个小鼠模型中证明了CD4+T细胞在肿瘤免疫中的直接作用。许多人类和小鼠肿瘤细胞上调MHC II类,尤其是在暴露于IFN-γ后,使CD4+T细胞能够通过颗粒酶直接进行细胞溶解。此外,小鼠和人类的CD4+T细胞也通过与吞噬细胞或B细胞协同作用,并通过协调免疫活动来驱动反应。
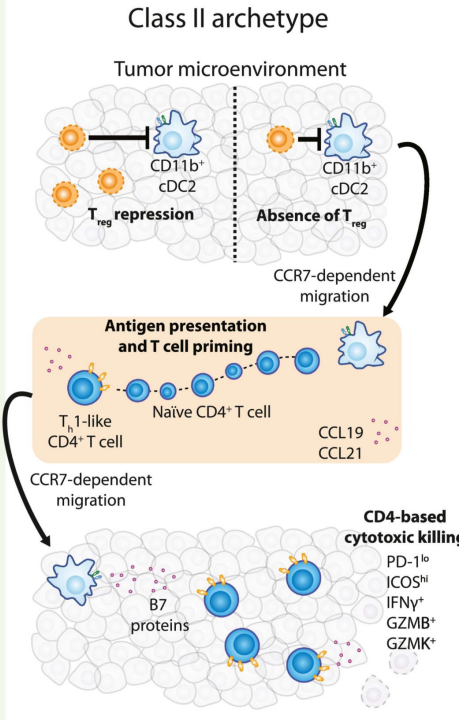
CD4+T细胞在II类反应性肿瘤免疫环境中的潜在伙伴通常是cDC2,在一项研究中,与其他髓系细胞表型相比,从荷瘤小鼠的TME中纯化的CD11b+cDC2在体外诱导CD4+T细胞扩增和增殖。在这项研究中,Treg细胞限制了cDC2深度刺激CD4+T细胞形成PD-1loICOS+表型并成为Teff细胞的能力。在人类中,头颈部肿瘤活检中发现cDC2和CD4+T细胞数量和表型之间存在相同的关系;此外,这些细胞频率和表型与Treg呈负相关。因此,在“阻断”的II类原型中,解除与Treg细胞招募、增殖和存活相关的通路可能有助于重建ICB反应性。
上调反应原型的途径
这些原型还有哪些其他特征有待揭示?下面是一些可能促进I类和/或II类反应原型的其他内在和外在因素。
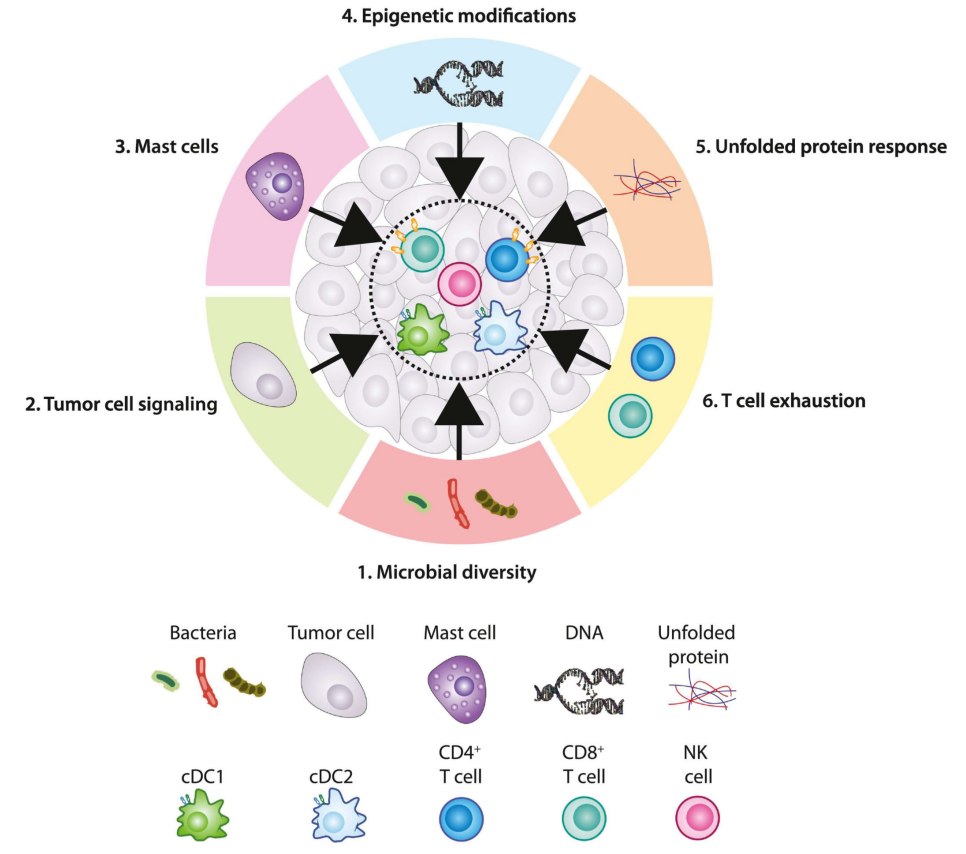
逆转T细胞耗竭
如前所述,肿瘤特异性TRM细胞和逆转“耗竭”T细胞可以增强免疫原型反应性。在一项关于人源化膀胱癌小鼠模型的研究中,表达CD40的cDC1通过逆转CD8+PD-1+LAG3+表型TEX信号诱导抗肿瘤免疫,并且与对照组相比,可以减少抗CD40抗体治疗小鼠的肿瘤生长。在另一个例子中,在黑色素瘤患者肿瘤中富集了五种不同的TEX表型,并显示出针对黑色素瘤相关抗原或新抗原的抗肿瘤活性。这些TEX细胞亚群,可能已经(i)重新激活——它们没有“完全”耗竭,保留了细胞溶解能力,(ii)激活了Teff记忆相关细胞亚群(TCF7-、表达IL7R),或(iii)激活LNs中产生的新T细胞。
表现遗传修饰
表观遗传学可以稳定TEX细胞,防止细胞再生,因此可能是ICB反应性的决定因素。通过深度测序(ATAC-seq)分析,发现TEX细胞的表观遗传学特征不同于肺癌患者的CD8+Teff和CD8+记忆T细胞。对人类肿瘤活检和荷瘤小鼠模型的进一步研究表明,CD8+T细胞分化程序中顺式(例如增强子/启动子)和反式(例如转录因子)中的基因调控元件可以改变TEX细胞重新编程以实现ICB反应的能力。因此,了解这些TEX表型如何塑造TME以及与其他免疫细胞类型的相互作用,这将是开发更好的策略以实现ICB反应性的关键。
调节内质网应激
内质网应激和未折叠蛋白反应(UPR)的激活有助于许多癌症的发生和进展。例如,DC上的XBP1和C/EBP同源蛋白(Chop)可以决定肿瘤中T细胞的发育、存活和激活。在卵巢癌荷瘤小鼠模型中,DC特异性XBP1缺失可抑制肿瘤生长,并增加CD4+和CD8+TIL上的IFN-γ表达。鉴于内质网应激与UPR激活之间的关系,阻断UPR可能被视为上调ICB反应性免疫原型的策略。
肥大细胞
肥大细胞(MCs)具有广泛的受体和共刺激分子,以快速响应传入信号,并分泌各种储存和新合成的介质。例如,MCs可以在人类和小鼠中产生Flt3L,并可能在I类反应性免疫原型中替代NK细胞。事实上,在非小细胞肺癌患者的肿瘤中发现的MC在应激状态下表达多种细胞因子,如TNF-α、IFN-γ、IL-6和糜蛋白酶,它们与这些患者的肿瘤生长抑制和OS改善相关。然而,在子宫内膜癌患者中,MC丰度与OS呈负相关,表明MC可能在不同癌症类型中表现出不同的功能。因此,有必要对MCs表型变异、结构和空间分布进行进一步研究,以了解和验证MCs如何以及在哪些表型中发挥反应性抗肿瘤或促肿瘤作用。
微生物组
小鼠研究表明,肠道细菌分类群与ICB反应之间存在关联。在一个研究中,健康的人类粪便微生物群(FMT)被移植到无菌小鼠体内,导致CD8+IFN-γ+T细胞扩增,并相对于对照组增强ICB介导的抗肿瘤免疫。在另一个研究中,非小细胞肺癌患者在存在嗜粘液阿克曼菌的情况下,血液中的CD4+Th1反应性增加;当比较ICB反应性和无反应性患者时,这与临床结果的改善相关。这些结果表明,在ICB治疗期间,微生物群可以影响I类和II类反应性免疫原型的关键成分。
小结
大量证据表明,ICB反应性至少有两种“类型”——I类和II类反应性免疫原型,这些原型的组成部分已经具有明确的预后,这些特征的结合有助于更好地预测ICB的反应性。对反应性原型的更全面理解也有可能调整临床前筛查平台,或与其他方法相结合以提高肿瘤治愈率。
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言



