
综述|肉毒毒素治疗局灶型肌张力障碍中枢神经系统机制的影像学研究进展
2024-11-27 中国神经精神疾病杂志 中国神经精神疾病杂志

文章揭示了BoNT可通过影响皮质、基底神经节、丘脑、小脑等脑区的功能连接、微观结构、代谢水平,以改善肌张力障碍患者的症状,为进一步探索肌张力障碍的发生机制和研发潜在的干预靶点提供理论参考。
摘 要 肉毒毒素(botulinum toxin, BoNT)是目前治疗局灶型肌张力障碍的一线方法,其机制为化学性去神经支配引起肌肉麻痹。近期神经影像学研究发现,BoNT亦可影响肌张力障碍患者脑内的神经可塑性,但具体的中枢神经系统机制尚未完全阐明。本文从功能磁共振成像、结构磁共振成像、正电子发射断层成像3个方面,对BoNT可能的中枢神经系统作用机制进行综述,揭示了BoNT可通过影响皮质、基底神经节、丘脑、小脑等脑区的功能连接、微观结构、代谢水平,以改善肌张力障碍患者的症状,为进一步探索肌张力障碍的发生机制和研发潜在的干预靶点提供理论参考。
关键词
肌张力障碍;肉毒杆菌毒素;中枢神经系统;神经网络;神经可塑性;磁共振成像;正电子发射断层成像
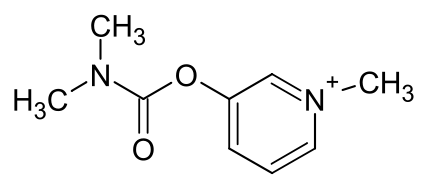
肌张力障碍是一种运动障碍疾病,特征是肌肉持续性或间断性收缩导致扭曲和重复的异常动作和(或)姿势异常[1]。按照症状分布,肌张力障碍可分为局灶型、节段型、多灶型、偏身型和全身型5种。其中局灶型肌张力障碍在临床中最为常见,包括颈部肌张力障碍(cervical dystonia, CD)、眼睑痉挛(blepharospasm, BSP)、书写痉挛、喉部肌张力障碍和口下颌肌张力障碍等[2-3]。目前,注射肉毒毒素(botulinum toxin, BoNT)是治疗局灶型肌张力障碍的一线方法[4],BoNT通过化学性去神经支配效应麻痹肌肉发挥作用,疗效一般为3~6个月。然而,临床实践发现,一些患者临床获益的时间远超过BoNT理论上肌肉麻痹时间。研究发现,BoNT可能通过外周神经逆向轴突转运和跨突触效应作用到中枢神经系统,从而影响肌张力障碍患者脑内的神经可塑性[5-8]。然而,BoNT治疗肌张力障碍的具体中枢神经系统机制尚未完全阐明。明确此机制将对肌张力障碍发生的研究和潜在干预靶点的研发具有重要意义。为此,本文将从功能磁共振成像、结构磁共振成像、正电子发射断层成像3个方面,对BoNT治疗肌张力障碍中枢神经系统机制的神经影像研究进展作以综述。
1 功能磁共振成像
1.1 静息态fMRI 静息态功能磁共振成像(resting state functional MRI, rs-fMRI)通常用于评估静息状态下受试者大脑的激活状态,同时可以避免其他混杂因素的影响,能全面反映BoNT治疗前后肌张力障碍患者的大脑激活状态改变,具有无创伤和较高的空间和时间分辨率等优点[9]。早期认为肌张力障碍是基底神经节功能障碍引起的,随着rs-fMRI技术迅速发展,研究表明肌张力障碍可能是一种“神经网络性疾病”,可能涉及皮质-基底神经节/小脑-丘脑-皮质等环路的功能障碍[10]。既往rs-fMRI研究表明BoNT治疗肌张力障碍患者诱导的神经可塑性改变可能涉及皮质、基底神经节、小脑等脑区。
感觉运动整合和高阶运动规划异常是局灶型肌张力障碍发生的重要原因。DELNOOZ等[11]应用rs-fMRI技术分析23例CD患者和22名健康对照的大脑网络,发现BoNT治疗前CD患者感觉运动网络中的前额叶皮质、运动前皮质(premotor cortex, PMC)和顶上小叶连接性降低,而治疗后感觉运动网络中的左侧额中回连接性增加、初级视觉网络中的双侧视觉皮质和初级运动皮质(M1)连接性增强。FENG等[12]对33例CD患者和45名健康对照进行rs-fMRI检查,应用体素级全局功能连接、局部一致性和基于种子点的功能连接分析等方法发现,与健康对照组相比,CD患者组右侧额上回和感觉运动皮质之间的功能连接增加,而BoNT治疗可减少这种异常的功能连接。以上两项研究结果提示,BoNT治疗可部分恢复CD患者功能连接受损的感觉运动网络,证明了额叶-感觉运动网络通路与BoNT治疗诱导的神经可塑性密切相关。但与上述研究不一致的是,MOHAMMADI等[13]使用rs-fMRI对16例书写痉挛患者和16名健康对照进行研究,发现与健康对照组相比,书写痉挛患者组的感觉运动网络和默认模式网络在功能连接上存在差异,但未发现书写痉挛患者在BoNT治疗前后的脑网络功能连接发生改变,提示BoNT治疗并未改变书写痉挛患者的潜在皮质功能障碍。
除了作用于感觉运动网络,BoNT的治疗作用还与基底神经节等运动控制及规划脑区密切相关。DELNOOZ等[14]应用rs-fMRI技术,分析BoNT治疗对23例CD患者基底神经节功能连接的影响,发现与BoNT治疗前相比,治疗后患者右腹中侧纹状体与执行控制网络之间功能连接增加,右外侧基底神经节与执行控制网络之间功能连接增加,提示经BoNT治疗后CD患者的脑网络功能连接重组。
近期,诸多神经病理学和神经影像学研究表明小脑在肌张力障碍的发病机制中亦起到关键作用[15-19]。HOK等[20]对17例CD患者进行rs-fMRI扫描发现,CD患者右小脑叶VⅥ、右小脑Crus Ⅱ区、蚓部小叶Ⅷb和右小脑叶Ⅸ的功能连接下降程度与临床症状改善程度呈正相关,BoNT治疗可改善小脑蚓部小叶Ⅷa与左背内侧额叶皮质之间的功能连接,恢复患者小脑部分区域的功能连接。另一项基于感兴趣区域功能连接的rs-fMRI研究,通过分析接受BoNT治疗的13例BSP/Meige综合征患者,发现BoNT治疗前患者小脑与躯体感觉皮质和视觉皮质之间的功能连接降低,而治疗后可部分恢复以上脑区之间的异常功能连接,提示BoNT可以通过改变BSP患者的本体感觉输入,改善其临床症状[21]。
以上研究fMRI扫描的时间节点均选在BoNT治疗后2~4周,集中分析了BoNT治疗高峰时间节点对大脑的即时调节作用。而BoNT改变肌张力障碍患者神经可塑性的rs-fMRI纵向研究相对较少。目前仅有一项针对111例接受BoNT治疗不同年限的喉部肌张力障碍患者和50名健康对照的大规模rs-fMRI纵向研究,结果发现,与较短(1~5年)或较长BoNT治疗年限(13~28年)患者相比,中间治疗年限(6~12年)的患者右侧小脑活动减少,且中间治疗年限(6~12年)患者额中回活动程度比较短治疗年限(1~5年)的患者减少[22]。这些结果提示,中间治疗年限的喉部肌张力障碍患者可从BoNT注射获得更多神经调节益处,这也间接解释了一部分患者随着BoNT治疗次数增加其临床获益逐渐减低甚至无获益[22]。
总之,以上rs-fMRI各项研究,可能由于临床异质性较大、研究方法差异、研究队列样本量小等,出现可重复性低甚至相互矛盾的研究结果。尽管如此,上述研究结论提示,BoNT治疗可部分恢复局灶型肌张力障碍患者的异常脑网络连接,主要集中在运动控制相关的各个脑区,如M1、顶叶、额叶、基底神经节、小脑等。
1.2 任务态fMRI 任务态fMRI(task fMRI, t-fMRI)是一种无创、无辐射的神经影像学方法,具有较强的针对性,可观察到不同任务刺激对受试者相应脑区的激活情况。
1.2.1 感觉任务激活fMRI 一项针对14例接受BoNT治疗的内收型痉挛性构音障碍患者和15名健康对照的t-fMRI研究发现,与健康对照组相比,治疗前触觉刺激患者的左手/面部和右面部可激活对侧初级体感皮质(S1),触觉刺激右面部可激活左侧次级体感皮质(S2)和颞上回,而BoNT治疗对S1、S2的激活程度无影响[23]。然而,BRODOEHL等[24]对17例经BoNT治疗的CD患者行rs-fMRI和对食指吹气的t-fMRI扫描发现,与健康对照相比,治疗前CD患者S1与S2功能连接减少,而基底节内部、基底节与丘脑、躯体感觉皮质与运动皮质、丘脑与壳核及丘脑与尾状核之间的功能连接增加。该研究还发现与治疗前相比,治疗后CD患者的S1与S2之间功能连接增加,而基底节内以及基底节、丘脑和感觉运动皮质之间的连接性降低,提示BoNT可能通过恢复感觉运动整合障碍影响CD患者的皮质可塑性。
虽然存在一些矛盾的研究结论,但以上研究仍然提示S1、S2感觉整合异常可能是几种肌张力障碍共同的病理生理学机制,而BoNT治疗可部分恢复局灶型肌张力障碍患者S1与S2区的激活异常。
1.2.2 运动任务激活fMRI NEVRLÝ等[25]在12例接受BoNT治疗的CD患者和12名健康对照执行手指运动任务时进行fMRI检查发现,与健康对照组相比,治疗前CD患者双侧辅助运动区(supplementary motor area, SMA)、同侧扣带回和副扣带回,以及同侧尾状核、苍白球和丘脑的激活程度明显降低。而治疗后CD患者与健康对照相比激活程度无差别。这些结果表明BoNT治疗后肌张力障碍患者皮质功能异常趋于正常化,提示改变肌张力障碍患者的本体感觉传入可能改变其中枢神经可塑性。
OPAVSKÝ等[26]对7例CD患者和9名健康对照在4 Hz正中神经电刺激时进行fMRI扫描,发现与健康对照相比,BoNT治疗前CD患者对侧S2、岛叶激活降低。而与BoNT治疗前相比,治疗后CD患者对侧S2、岛叶、顶下小叶激活增加,提示BoNT治疗可以使肌张力障碍患者躯体感觉整合障碍部分正常化。此外,HASLINGER等[27]使12例喉部肌张力障碍患者执行发声任务并进行fMRI扫描发现,在BoNT治疗前,发声任务诱发患者初级感觉运动皮质、PMC的激活减少,但在BoNT治疗后血氧水平依赖信号并无差别,提示BoNT治疗不能逆转喉部肌张力障碍患者神经可塑性的异常改变。
由于以上t-fMRI研究中使用的任务方式不同,同时研究方法、参数亦有所差异,在细微的影像学改变识别、激活脑区和激活程度等方面难以得到一致的研究结论。总之,t-fMRI研究结果显示BoNT治疗对肌张力障碍患者的中枢神经系统影响范围较广,主要集中在基底节区、丘脑、运动皮质等部位,间接证明了局灶型肌张力障碍是一种全脑神经网络障碍性疾病。
2 结构磁共振成像
结构磁共振成像可通过定量测量大脑皮质厚度、灰质体积、白质密度等影像学指标,识别大脑微观结构的改变,为BoNT治疗肌张力障碍患者的中枢神经系统机制提供客观的影像学证据。
有研究利用结构性MRI结合基于体素的形态学测量方法发现:与偏侧面肌痉挛患者相比,BSP患者额叶和颞叶皮质(包括感觉运动皮质)的连接性增加,小脑的连接性降低;与治疗前相比,BoNT治疗后BSP患者左侧感觉运动皮质连接性降低,但治疗后偏侧面肌痉挛患者的小脑连通性下降[28]。该研究结果提示,BoNT治疗偏侧面肌痉挛患者的症状改善与小脑连通性下降相关,BoNT对BSP患者小脑等部位的潜在皮质功能障碍无显著影响。BLOOD等[29]利用弥散张量成像技术对接受BoNT治疗的14例CD患者进行分析,发现治疗前患者左侧大脑半球的部分各向异性指数(fractional anisotropy, FA)降低,而治疗后患者左侧大脑半球FA增加。另外,BoNT治疗4周后Tusi评分下降与不对称FA值改变呈正相关,提示BoNT治疗有效改善运动症状,同时还改变了CD患者白质的微观结构。未来神经影像学研究不仅需探索大脑半球之间的白质变化,还可进一步探索BoNT治疗对基底神经节、丘脑、小脑等局部白质纤维束的影响。
3 正电子发射断层成像
正电子发射断层成像(positron emission tomography, PET)是基于局部示踪剂浓度检测脑组织活性强度的分子成像技术,可根据不同示踪剂在不同组织中的聚集浓度评估脑组织功能[30]。
一项18F-脱氧葡萄糖PET研究,探索清醒状态BSP患者接受BoNT治疗后的脑代谢状态变化。根据BoNT治疗后的疗效不同,将25例BSP患者分为完全改善组和不完全改善组,发现与健康对照组相比,不完全改善组的双侧丘脑、脑桥和小脑代谢增高;与健康对照组相比,完全改善组则仅在双侧丘脑中观察到高代谢,提示丘脑代谢增高可能是BSP发生的关键病理生理变化[31]。然而该研究并没有分析BoNT治疗前BSP患者的脑代谢情况,未进行治疗前后脑代谢的对比,存在一定的局限性。
4 总结与展望
综上所述,神经影像学研究发现局灶型肌张力障碍的发生机制可能与感觉运动皮质、丘脑、小脑、基底节等多个脑区的结构、功能连接、代谢状况发生改变密切相关,BoNT治疗可部分恢复这些脑区的异常激活或异常连接。这些结果提示肌张力障碍不是由单一节点障碍引起的,进一步验证了全脑神经网络功能紊乱可能是肌张力障碍发生的重要机制。同时,BoNT治疗肌张力障碍的神经影像学研究可能为神经调控治疗,如脑深部刺激、经颅磁刺激或经颅直流电刺激等[32-33],提供新的干预靶点。例如,BoNT治疗可部分恢复患者特定脑区(如SMA、PMC、M1、S1、S2、岛叶、顶叶、基底节、丘脑、小脑等)的异常激活或异常连接,提示上述脑区可作为神经调控技术治疗肌张力障碍的参考靶区。因此,未来临床研究可聚焦新的治疗靶区,为肌张力障碍治疗带来新的希望。然而,当前的研究在一定程度上均存在一些局限性,如临床异质性较大、样本量小、纵向研究缺乏、先进影像学技术和分析方法应用相对匮乏等。未来应开展大样本、多中心、多模态MRI的纵向研究,帮助进一步阐明BoNT治疗肌张力障碍的中枢神经系统机制,为实现精准治疗提供更加可靠、客观的临床依据。
参考文献:
1. BALINT B, MENCACCI N E, VALENTE E M, et al. Dystonia[J]. Nat Rev Dis Primers, 2018, 4(1): 25.
2. TEIVE H A G, CHEN C C. Isolated focal dystonia: The mysterious pathophysiology is being unraveled[J]. Neurology, 2020, 95(16): 711-712.
3. 穆宇辰, 苏俊辉, 滕飞, 等. 肌张力障碍缓解技巧现象研究进展[J]. 中国神经精神疾病杂志, 2020, 46(9): 558-561.
4. THENGANATT M A, JANKOVIC J. Treatment of dystonia[J]. Neurotherapeutics, 2014, 11(1): 139-152.
5. MATAK I. Evidence for central antispastic effect of botulinum toxin type A[J]. Br J Pharmacol, 2020, 177(1): 65-76.
6. ŠOŠTARIĆ P, VUKIĆ B, TOMAŠIĆ L, et al. Lasting peripheral and central effects of botulinum toxin type A on experimental muscle hypertonia in rats[J]. Int J Mol Sci, 2022, 23(19): 11626.
7. ANTONUCCI F, ROSSI C, GIANFRANCESCHI L, et al. Long-distance retrograde effects of botulinum neurotoxin A[J]. J Neurosci, 2008, 28(14): 3689-3696.
8. CALEO M, SPINELLI M, COLOSIMO F, et al. Transynaptic action of botulinum neurotoxin type A at central cholinergic boutons[J]. J Neurosci, 2018, 38(48): 10329-10337.
9. FINN E S, POLDRACK R A, SHINE J M. Functional neuroimaging as a catalyst for integrated neuroscience[J]. Nature, 2023, 623(7986): 263-273.
10. THOMSEN M, LANGE L M, ZECH M, et al. Genetics and pathogenesis of dystonia[J]. Annu Rev Pathol, 2024, 19(1): 99-131.
11. DELNOOZ C C S, PASMAN J W, BECKMANN C F, et al. Task-free functional MRI in cervical dystonia reveals multi-network changes that partially normalize with botulinum toxin[J]. PLoS One, 2013, 8(5): e62877.
12. FENG L, YIN D, WANG X, et al. Brain connectivity abnormalities and treatment-induced restorations in patients with cervical dystonia[J]. Eur J Neurol, 2021, 28(5): 1537-1547.
13. MOHAMMADI B, KOLLEWE K, SAMII A, et al. Changes in resting‐state brain networks in writer’s cramp[J]. Hum Brain Mapp, 2012, 33(4): 840-848.
14. DELNOOZ C C S, PASMAN J W, BECKMANN C F, et al. Altered striatal and pallidal connectivity in cervical dystonia[J]. Brain Struct Funct, 2015, 220(1): 513-523.
15. LI H, ZHANG M, WU Y, et al. Iron accumulation and axonal damage of cerebellum in idiopathic cervical dystonia[J]. Eur J Neurol, 2023, 30(6): 1619-1630.
16. MACIVER C L, BAILEY G, LAGUNA P L, et al. Macro- and micro-structural insights into primary dystonia: A UK Biobank study[J]. J Neurol, 2024, 271(3): 1416-1427.
17. CORP D T, JOUTSA J, DARBY R R, et al. Network localization of cervical dystonia based on causal brain lesions[J]. Brain, 2019, 142(6): 1660-1674.
18. ZITO G A, TARRANO C, JEGATHEESAN P, et al. Somatotopy of cervical dystonia in motor-cerebellar networks: Evidence from resting state fMRI[J]. Parkinsonism Relat Disord, 2022, 94: 30-36.
19. FAGAN M, SCORR L, BERNHARDT D, et al. Neuropathology of blepharospasm[J]. Exp Neurol, 2021, 346: 113855.
20. HOK P, HVIZDOŠOVÁ L, OTRUBA P, et al. Botulinum toxin injection changes resting state cerebellar connectivity in cervical dystonia[J]. Sci Rep, 2021, 11: 8322.
21. JOCHIM A, LI Y, GORA‐STAHLBERG G, et al. Altered functional connectivity in blepharospasm/orofacial dystonia[J]. Brain Behav, 2018, 8(1): e00894.
22. O’FLYNN L C, SIMONYAN K. Short- and long-term central action of botulinum neurotoxin treatment in laryngeal dystonia[J]. Neurology, 2022, 99(11): e1178-e1190.
23. MANTEL T, DRESEL C, WELTE M, et al. Altered sensory system activity and connectivity patterns in adductor spasmodic dysphonia[J]. Sci Rep, 2020, 10: 10179.
24. BRODOEHL S, WAGNER F, PRELL T, et al. Cause or effect: Altered brain and network activity in cervical dystonia is partially normalized by botulinum toxin treatment[J]. Neuroimage Clin, 2019, 22: 101792.
25. NEVRLÝ M, HLUŠTÍK P, HOK P, et al. Changes in sensorimotor network activation after botulinum toxin type A injections in patients with cervical dystonia: A functional MRI study[J]. Exp Brain Res, 2018, 236(10): 2627-2637.
26. OPAVSKÝ R, HLUŠTÍK P, OTRUBA P, et al. Somatosensory cortical activation in cervical dystonia and its modulation with botulinum toxin: An fMRI study[J]. Int J Neurosci, 2012, 122(1):45-52.
27. HASLINGER B, ERHARD P, DRESEL C, et al. “Silent event-related” fMRI reveals reduced sensorimotor activation in laryngeal dystonia[J]. Neurology, 2005, 65(10): 1562-1569.
28. CHIRUMAMILLA V C, DRESEL C, KOIRALA N, et al. Structural brain network fingerprints of focal dystonia[J]. Ther Adv Neurol Disord, 2019, 12: 175628641988066.
29. BLOOD A J, KUSTER J K, WAUGH J L, et al. White matter changes in cervical dystonia relate to clinical effectiveness of botulinum toxin treatment[J]. Front Neurol, 2019, 10: 265.
30. MILLEVERT C, VIDAS-GUSCIC N, VANHERP L, et al. Resting-state functional MRI and PET imaging as noninvasive tools to study (Ab)normal neurodevelopment in humans and rodents[J]. J Neurosci, 2023, 43(49): 8275-8293.
31. SUZUKI Y, MIZOGUCHI S, KIYOSAWA M, et al. Glucose hypermetabolism in the thalamus of patients with essential blepharospasm[J]. J Neurol, 2007, 254(7): 890-896.
32. BLEDSOE I O, VISER A C, SAN LUCIANO M. Treatment of dystonia: Medications, neurotoxins, neuromodulation, and rehabilitation[J]. Neurotherapeutics, 2020, 17(4): 1622-1644.
33. KRAUSS J K, LIPSMAN N, AZIZ T, et al. Technology of deep brain stimulation: Current status and future directions[J]. Nat Rev Neurol, 2021, 17(2): 75-87.
【引用格式】穆秦颖,胡俊豪,马福浩,等.肉毒毒素治疗局灶型肌张力障碍中枢神经系统机制的影像学研究进展[J]. 中国神经精神疾病杂志,2024,50(7):440-444.
【Cite this article】MU Q Y,HU J H,MA F H,et al. A progress in neuroimaging research on the central mechanisms of botulinum toxin in the treatment of focal dystonia[J]. Chin J Nervous Mental Dis,2024,50(7):440-444.
DOI:10.3969/j.issn.1002-0152.2024.07.008
本网站所有内容来源注明为“梅斯医学”或“MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源为“梅斯医学”。其它来源的文章系转载文章,或“梅斯号”自媒体发布的文章,仅系出于传递更多信息之目的,本站仅负责审核内容合规,其内容不代表本站立场,本站不负责内容的准确性和版权。如果存在侵权、或不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言











#肉毒杆菌毒素# #中枢神经系统# #肌张力障碍#
19